先日お伝えした太陽光パネル問題に続いて、今度は風力発電が問題になっている。巨大風力発電設備が低周波による人への健康被害や自然の生態系にも影響を及ぼしている。風力発電は太陽光と違ってまだリスクが周知されていない。具体的には睡眠障害、睡眠遮断、耳鳴り、頭痛、耳鳴り、めまい、吐き気、集中力や記憶力の異常。低周波音や超低周波音は遮音壁やガラスでは防げない。扇風機が回転する低周波音は床に耳を付けていると分かる。
また低周波音を嫌ってクマやイノシシが麓に出現して凶暴化するとの報告も。国の規制は無いに等しく、経産省や環境省の許可制ではなく認証に過ぎないため、市民からの意見を採用するかどうかは事業者次第。風力発電のメリットは太陽光と同じFIT:固定価格買い取り制度とFIP:利益保証型買い取り制度でボロもうけ状態。これらの元となるのは国民が支払う再エネ賦課金である。
![]()
![]()
![]()
![]()
![]()
荒川央先生:RNAコロナワクチンにDNAが混入している?

コロナワクチン接種者の体内で数ヶ月以上の期間、スパイクタンパクが血中を循環する事が報告されています。なぜこれほど長い間スパイクタンパクが体内に残るのか? 体内でスパイクタンパクの生産が続いているのではないか? また、シュードウリジン化RNAが安定であるとしても、何ヶ月以上も安定に存在し得るのか? 数多くの疑問が浮かんできます。
厚生労働省は「新型コロナワクチンQ&A」において以下のように記しています。
mRNA (メッセンジャーRNA) ワクチンで注射するmRNAは、数分から数日といった時間の経過とともに分解されていきます。また、mRNAは、人の遺伝情報 (DNA) に組みこまれるものではありません。身体の中で、人の遺伝情報 (DNA) からmRNAがつくられる仕組みがありますが、情報の流れは一方通行で、逆にmRNAからはDNAはつくられません。こうしたことから、mRNAを注射することで、その情報が長期に残ったり、精子や卵子の遺伝情報に取り込まれることはないと考えられています。
この短い文章の中にも既にいくつもの誤りが見られます。ワクチンで作られるmRNAは、シュードウリジンのためにRNA分解経路に対して耐性であり、実際に胚中心内で少なくとも2ヶ月もの間残っている事が確認されています。逆転写の仕組みはmRNAをDNAに変換できますし、ヒトのゲノム内にもレトロポゾン由来の逆転写酵素が存在します。
コロナウイルスのゲノムはRNAであり、RNAワクチンの遺伝情報もRNAです。ヒトのゲノムはDNAなので、ヒトゲノムにスパイクタンパクの遺伝子が取り込まれるためには、その遺伝情報がDNAである必要があります。私のブログでもRNAワクチンの逆転写については何度か取り上げてきました。しかし、コロナワクチンにスパイクタンパクDNAが含まれている場合には、ゲノムへの取り込みに逆転写すら必要条件ではなくなるのです。
アストラゼネカ、ジョンソン&ジョンソンのコロナワクチンはアデノウイルスベクターによるDNAワクチンです。アデノウイルスベクターは遺伝子治療などにも応用されていますが、ゲノムに取り込まれる事もあります。おそらくDNAワクチン接種者の中にはワクチンのDNAをゲノムに取り込み、恒久的にスパイクタンパク遺伝子を発現している人が既に存在しているでしょう。
例えば、生まれつき欠損している遺伝子の機能を「補う」ために行われる遺伝子治療の場合には、その遺伝子がゲノムに取り込まれたとしても問題は限定的です。しかし、スパイクタンパクのような毒性の高い遺伝子では話は違ってきます。
ファイザー、モデルナのRNAワクチンに含まれるRNAは、その鋳型となるDNAから転写して作られたものです。転写の鋳型となったDNAがRNAワクチンに混入しているのではないかという疑惑が現在持たれています。発端はMedicinal Genomics社のKevin Mckernan博士のブログ上での報告です。博士は二価コロナワクチンをディープシークエンシングした結果、ワクチンの中にプラスミドDNAの混入を発見しました。
Mckernan博士の当初の目的は、RNAコロナワクチンの遺伝子品質のチェックでした。RNAワクチンには細胞内免疫系を回避するために1メチルシュードウリジンが使われています。そして、ワクチン内のRNAは鋳型DNAからRNAを転写して作られています。1メチルシュードウリジンはこの転写の際にRNAに取り込まれますが、その際にエラーが起こりやすいのです (転写エラー率は4000ヌクレオチドあたり1エラー)。
ファイザーやモデルナのRNAワクチンのサイズは約4000ヌクレオチドですので、つまり、合成されたほとんどのワクチンRNA分子にエラーがあってもおかしくないという事になります。エラー率はスパイクタンパクの品質管理において重要な情報です。そして、シュードウリジンがタンパクへの翻訳に与える影響は未知数です。転写と翻訳のエラー率を考えると、RNAワクチンから多様な異常スパイクタンパクが作られる可能性が否定できません。こうした異常スパイクタンパクの中には自己免疫疾患やプリオン病の原因となるものも含まれるかもしれません。
この実験はそうしたエラーを検定する事が当初の目的だったのですが、ディープシークエンシングの過程で想定していなかったものが出てきました。ワクチンRNA合成の元となったプラスミドDNAの混入です。プラスミドは、染色体とは独立して複製することができる染色体外DNA分子です。人工的に作られたプラスミドは遺伝子クローニングに利用できるため、生命科学や遺伝子工学の分野では広く応用されています。
DNAを動物細胞内へ導入する手法はトランスフェクションと呼ばれます。細胞への導入法も様々ですが、脂質粒子 (リポソーム) を用いたリポフェクション法も汎用される方法です。この手法は脂質ナノ粒子によるRNAワクチンの細胞への導入と似ています。本来、もしたとえDNAが細胞内に導入され、核へ侵入したとしても、必ずしもそのDNAがゲノムに組み込まれるわけではありません。
そのため、ゲノムへの組み込み率を上げるための手段としてはウイルスベクター (レンチウイルスベクターやレトロウイルスベクターなど) や、トランスポゾンベクターなどがあり、また最近ではCRISPRを用いたゲノム編集もよく使われます。しかし、そうした新しい技術を用いなくともDNAを細胞に導入すると、効率は低くなりますが、ゲノムにも取り込まれるのです。
そうした方法は古典的な技術ではあるのですが、今でも汎用されています。「トランスジェニック」とは外来遺伝子の導入を意味する専門用語です。例えば、外来遺伝子をゲノムに組み込んだマウスはトランスジェニックマウスと呼ばれます。もしコロナワクチンにDNAが含まれるならば、ワクチンを接種しているだけのはずがDNAを人体にトランスフェクションしてしまった事になります。つまり、このDNAが人間のゲノムに組み込まれた場合、まさにトランスジェニック人間になるのです。
博士はコロナワクチンRNAの配列をディープシークエンシングする手法を使いました。この技術はRNA seq (アール・エヌ・エー・セク) と呼ばれます。この技術ではまずはRNAを逆転写してDNAにしてから配列を決定します。理由は、RNAのままで配列を決定したり、増幅したりするのが難しいからです。このように分子生物学の実験手法でRNAの塩基配列を解析したり、遺伝子を増幅したりする場合、まずはRNAをDNAに変換する必要があります。
例えば、コロナウイルス感染のPCR検査でも、コロナウイルスのRNAゲノムを逆転写してからPCR増幅をかけています。コロナの遺伝子が検出された場合、実際にはRNAから増幅されたものとDNAから増幅されたものは通常区別されません。RNA seqの実験でも、元から存在したDNAを分解していない場合、DNAも一緒に配列を決定してしまうのです。今回、DNAの混入はそうした過程で見つかりました。
RNA seqの結果から、混入したDNAの量はRNA量の1/3000と見積もられました 。しかし、これはもともとはRNAの配列を決めるための実験ですので、基本的にはDNAの検出に最適な条件でなされたものではありませんでした。そのため、DNAに焦点を当ててさらに再解析がなされ、複数の方法で混入したDNAの定量化と解析がなされました。定量的PCR、電気泳動、大腸菌への導入、DNAに集中してのディープシークエンシングなどです。
また、博士がモデルナのコロナワクチンの2つの異なるロットを解析したところ、モデルナの二価ワクチンには、2種類の発現ベクターが含まれていました。混入しているプラスミドベクターの配列は汎用されているあるプラスミドと99.8%同一の配列を含んでいました。
EMAは、1 mg RNA当たりの二重鎖DNA汚染の限界を330 ng未満に設定しています。これは、mRNA3030分子あたりおよそ1つの割合になります。この基準がどのように設定されたのかは不明です。そもそも「どの量以下のDNAならゲノムに取り込まれない」などの基準は存在しないからです。しかしながら、DNAに焦点を絞って解析した結果、汚染レベルは当初の見積もりよりも100倍高く、各ワクチンの核酸の20-35%が発現ベクターでした (DNAの混入量は8.19~11.3 ng/ulで、mRNAは23-55ng/ul)。これは、EMAの制限値である330 ng/mg RNAを数桁超えており、1回のワクチン接種で数兆個のDNA分子が投与される事を意味します。
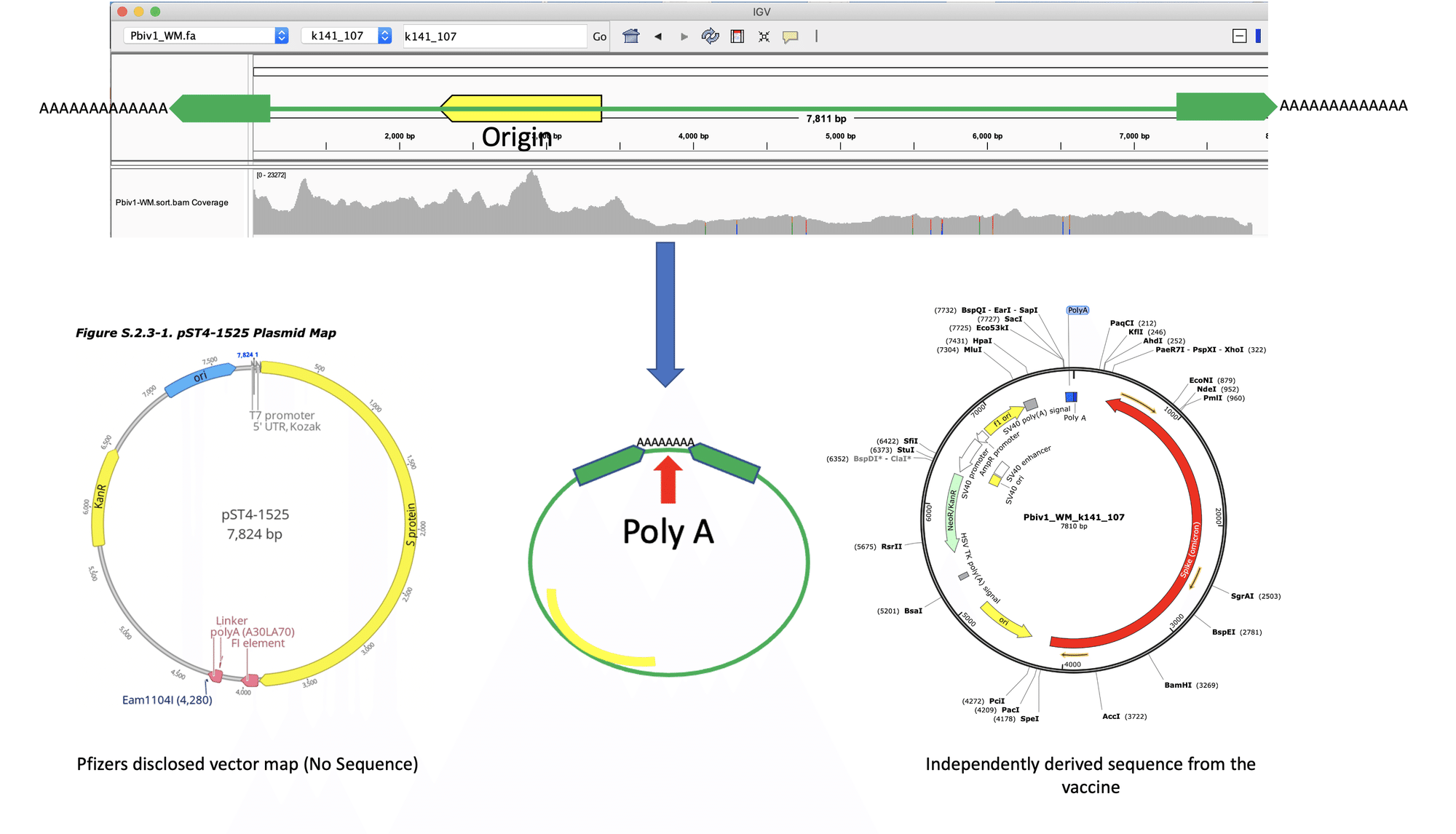
図の左側はファイザーが開示したベクターマップです。真ん中のプラスミドは、ディープシークエンシングの解析結果を描写したものです。参照した配列の両側にまたがるシークエンスリードがある事は、環状DNAが含まれる事を意味します。右側のベクターは、コロナワクチンに混入していたDNAの塩基配列から再現されたものです。検出されたDNAの塩基配列はファイザーが開示したものとほぼ同じでしたが、一部に違いが見られました。
DNAの形状は大きく分けて2種類あります。環状か直鎖状です。ディープシークエンシングによって分かったのは、ファイザー社の二価ワクチンに環状プラスミドが存在しているという事です。大腸菌内で増幅できるプラスミドベクターは環状DNAなのですが、環状プラスミドの存在は大腸菌への導入実験によっても裏付けられ、混入したDNAは菌内で直接増幅する事ができました。環状DNAと直鎖状DNAの正確な比率は現時点では不明です。環状プラスミドがゲノムに挿入される際には、DNAはどこかで切断され、それが組み込まれる事になります。ゲノムにDNAを導入する場合には、環状DNAを酵素処理して直鎖状に変換する手法もよく使われますが、それは重要な遺伝子や制御領域が偶発的に壊れないようにするためです。環状DNAがゲノムに組み込まれる場合、DNA内のどこで切断されて、どう組み込まれるかはランダムです。実際そのランダムさのために、同じDNAがゲノムに組み込まれても人によって起こりうる副作用にも多様性が生じると考えられます。
塩基配列の比較からさらに分かってきた事もあります。それは、ファイザー社の同じロットのワクチンから見つかったプラスミド内で、既知のSV40プロモーターの改良が見られた事です。SV40 (simian virus 40) はサルとヒトの両方に感染できるポリオーマウイルスです (「simian」は「サル」を意味します) 。癌の研究においてもSV40は重要な貢献をしてきました。例えば、重要な癌抑制遺伝子であるp53は最初、SV40のラージT抗原と結合するタンパクとして発見されたものです。SV40はDNAウイルスであり、染色体外で複製できるので、DNA複製のモデル系となっています。またSV40の強力なプロモーターは細胞への遺伝子導入と強制発現のために汎用されています。
特徴的な事として、同じロットのファイザーコロナワクチンの2つのバイアルから、2種類の異なる配列が得られました。2つのバイアルの混入DNAを比較すると、1つのバイアルのDNAにはSV40プロモーターにタンデム重複 (同一方向向きの繰り返し配列) があったのです。これはSV40プロモーターをターボ化する方法として知られています。ここから、混入したDNAの間に、こうした「改良」が行われている事が分かりました。
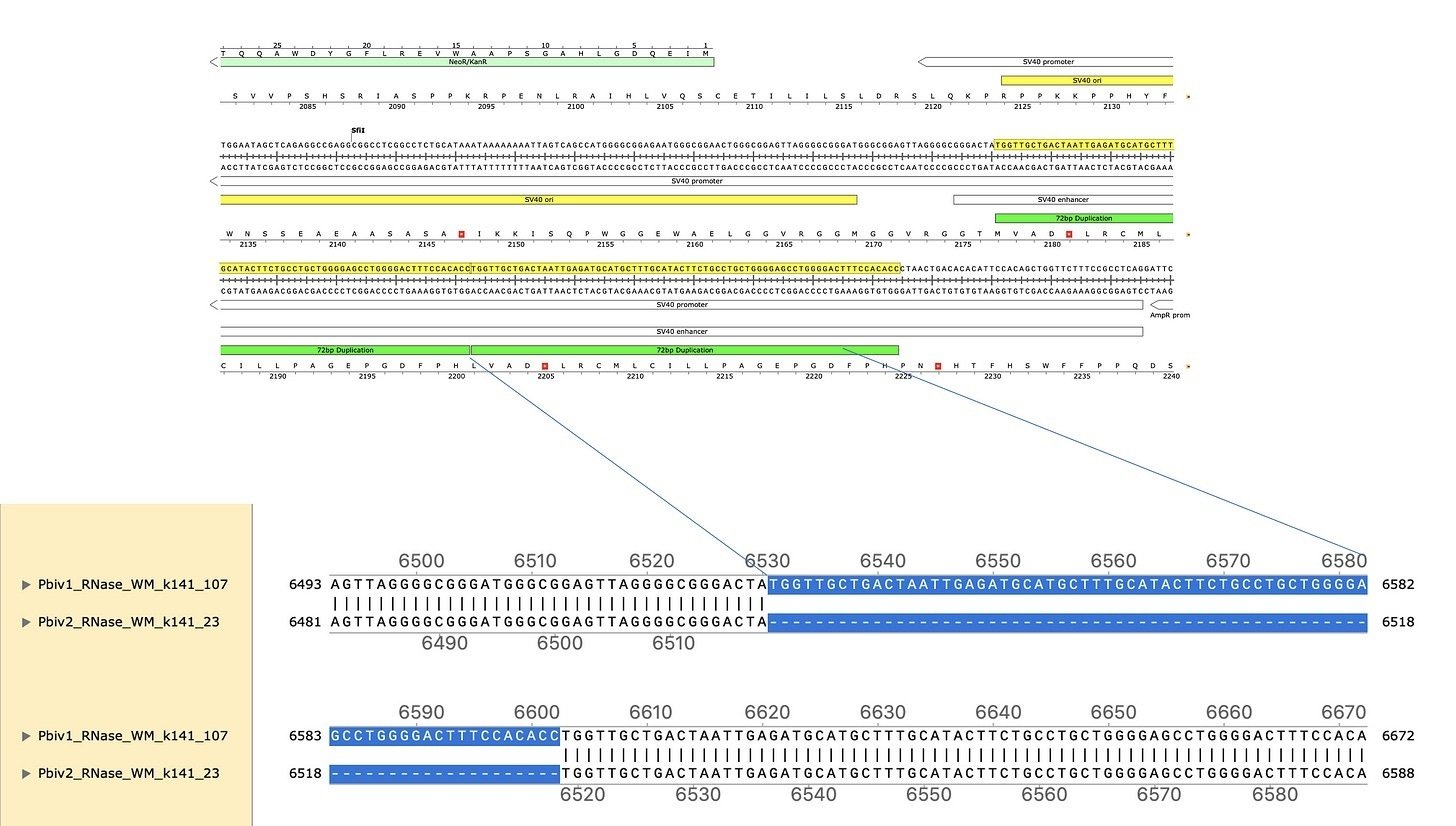
SV40ウイルスのゲノムは細胞核内で増幅します。その目的に適応するように、SV40プロモーターはDNAの核への取り込みを媒介する領域 (SV40プロモーター内のエンハンサー領域の72bp配列) を持っているのです。さらにこの配列を繰り返すと、核への輸送はさらに促進されます。
Sequence requirements for plasmid nuclear import
https://pubmed.ncbi.nlm.nih.gov/10585295/
博士はさらにファイザーの一価mRNAワクチンの同じロット (ロット# FL8095) から8本のバイアルを調査しました。どのバイアルにもDNAは一貫して混入しており、RNA:DNAの比率も同じロット内で一定でした。これはファイザー社の一価ワクチンにおけるDNAの混入もEMAの基準をはるかに超えていました。
RNAワクチンにDNAの汚染がある事自体、非常に大きな問題なのですが、不思議な点がいくつもあります。まずは、汚染レベルがなぜこれ程までに高いのかという事です。ワクチンのRNAはT7 RNAポリメラーゼによる転写によって作られるのですが、この酵素による転写は効率的で、その際のシュードウリジンのRNAの取り込みも高効率で起こります。ワクチンのRNAを作成するにしてもこれほど大量の鋳型DNAは不要なはずです。
鋳型DNAはDNA分解酵素などの処理で分解するはずであったと考えられますが、DNA分解酵素も効率が高いのです。これは、本当にそうした処理が不十分だっただけなのでしょうか?
さらに不思議な点は、不要なはずのSV40プロモーターがベクター上に存在する事です。この配列はプラスミドを核に輸送するので、ゲノムに取り込まれるリスクをさらに上げる事になります。そして、別のプラスミドではSV40プロモーターの繰り返し配列が見られるのです。このプラスミドの「改良」によって転写活性や核輸送がさらに促進されます。
では、こうしたDNAの混入はコロナワクチン後遺症にどう影響し得るでしょうか?
ここから先は私自身の考察も含む内容になります。
リスク1) 免疫反応の過剰刺激
外部から細胞内に取り込まれた二重鎖DNAは免疫系に感染体とみなされるため、自然免疫反応を強く刺激します (具体的には、STINGを介してI型インターフェロン反応を誘導します)。またプラスミドDNAは大腸菌から精製するものですが、精製の際にしばしばエンドトキシン (内毒素: グラム陰性菌の細胞壁の成分であるリポ多糖) が混入します。これらは炎症反応やアナフィラキシーの原因となります。
リスク2) 常在菌内でのDNA増幅
混入したDNAは菌内で増幅可能なプラスミドベクターである事です。コロナワクチン接種者の細菌に取り込まれると、DNAは細菌内で膨大な量に増幅される可能性があります。プラスミドベクターにはカナマイシン耐性遺伝子が組み込まれているので、接種者が抗生物質のカナマイシンを投与されている場合には、菌内でDNAが増幅するリスクはさらに上昇します。ただし、ヒトと菌での遺伝暗号の違いのために、菌内でスパイクタンパクへの翻訳は起こらないはずです。
リスク3) DNAの核への移行
ゲノムは細胞核内にあるので、DNAがゲノムに取り込まれるためには核へ移行する必要があります。そしてこのプラスミドDNAには核への移行を媒介する配列、つまりSV40プロモーターが存在するのです。
リスク4) スパイクタンパク遺伝子のゲノムへの取り込み
逆転写するまでもなく、コロナワクチンにはDNAの混入があったわけです。しかもそこには全長のスパイク遺伝子が含まれています。
では、ゲノムに取り込まれた場合にはどのような事態が想定されるでしょうか?
リスク5) ゲノムに取り込まれたスパイクタンパクが恒久的に発現する可能性
デザイン上はスパイクタンパクの遺伝子の発現にはファージ (菌に感染するウイルス) のプロモーターが使われており、ゲノムに取り込まれる事が直接スパイクタンパク遺伝子の発現につながるわけではありません。
しかし、ゲノムに取り込まれた位置によっては強力なプロモーターの近傍に配置される可能性があります。そうした場合、スパイクタンパクの生産はその後止むことなく続くわけです。時間とともに悪化していく多様なコロナワクチン後遺症に繋がり得ます。
リスク6) ゲノムに取り込まれた位置によって様々な遺伝子との融合タンパクが作られる可能性
リスク7) ファイザーのコロナワクチンの相補鎖 (二重鎖DNAの反対側) に存在する未知の遺伝子を発現する可能性
ブレーキの無いRNAワクチン
https://note.com/hiroshi_arakawa/n/n3111d6b3b0e0
リスク8) 癌
SV40プロモーターががん遺伝子の近傍に取り込まれれば癌遺伝子は恒常的に発現するようになります。細胞癌化の原因になります。こうしたものもワクチン接種後のターボ癌の一つの原因と考えられます。
コロナワクチンにスパイク遺伝子を含むDNAの混入があるのならば、それは大スキャンダルです。コロナワクチンが危険な理由の1つは、その成分の全てが公開されてはおらず、実際には何が入っているか分からないという点にもあります。コロナワクチンの毒性がスパイクタンパクのみで全て説明できるとも限らないという事です。