「“因果関係が否定できない”などとぼやかすのでなく、リスクや副反応については原因を究明して情報を出すべき」と厚労省の審査に注文を付けています。密室で行われている因果関係判定ですが、堂々と外に出して議論する姿勢は全くありません。重大なことを密室で決めてしまう、この姿勢は厳しく問われるべきです。
14歳の女子中学生が新型コロナウイルスの3回目のワクチン接種後に死亡し、医師らが接種と死亡との因果関係を認める結論を出しました。 徳島大学によりますと、去年8月に当時中学3年の14歳の女子中学生がファイザー製の新型コロナワクチンの3回目を接種しました。その翌日に37.9℃の発熱がありましたが、夕方には熱が下がったため、そのまま就寝しました。
しかし翌朝、心肺停止の状態で見つかり、その後、死亡が確認されました。女子生徒は、1回目の接種後は「発熱がなく腕の痛み」、2回目は「接種翌日に38℃未満の発熱」があったということです。 徳島県警は「健康な10代の女性が突然亡くなるのはおかしい」として、死因を調べるために徳島大学に司法解剖を依頼。解剖した医師らは、女子中学生に基礎疾患やウイルス感染などが無く本質的に健康で、中学の運動クラブで活動する生徒だったにもかかわらず、肺や肝臓など主要な臓器に炎症が起きて心不全になったことなどから、「ワクチン接種と死亡に因果関係あり」と結論付けたということです。
この結果は日本法医学会の医学誌「リーガルメディスン」で発表されました。論文をまとめた徳島大学法医学教室の主田英之准教授はMBSの取材に対して「今回は、司法解剖ができて、原因が特定できたことが大きい」と話しました。さらに主田准教授は「今回の1例でワクチン接種=危険とは判断できない」とも話し、国や医師に対して「“因果関係が否定できない”などとぼやかすのでなく、リスクや副反応については原因を究明して情報を出すべき」と求めました。
私たちに対しては「その情報を見て、接種について個人的・社会的な効果とリスクを総合的に考えて判断してほしい」と話しました。 ▼MBS NEWS HP
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
自己増殖型mRNAワクチンに関して警鐘が鳴らされています。
・生涯ワクチンを周囲に拡散する人が出現する可能性
・RNAワクチンの複製にはDNA修復機構がエラーを最小限に抑える修復機構がない
・突然変異で自己免疫疾患を誘発する可能性
・人体の中で「自己増殖型mRNAワクチンが進化」し、「増殖しやすいワクチン」や「免疫系を回避しやすいワクチン」、「感染しやすいワクチン」が派生する事
・遺伝子増殖にブレーキがなく、アクセルだけは付いており、再び感染拡大の恐れ
自己増殖型mRNAワクチン

今回は、近頃SNS上等でも話題になっている自己増殖型mRNAワクチンについてお話ししようと思います。
セントラルドグマはフランシス・クリックが1958年に提唱した分子生物学の概念で、遺伝情報が「DNA -> mRNA -> タンパク質」の順に伝達されるというものです。本来、RNAからRNAをコピーする仕組みをヒト細胞は持っていません。セントラルドグマには例外が2つ見つかっています。一つはレトロウイルスで見つかった逆転写です。この反応は逆転写酵素によってRNAを鋳型としてDNAを合成 (逆転写) するものです。そして、もう一つの例外がRNAウイルスから見つかったRNA依存性RNA複製です。この仕組みにおいて、RNA依存性RNAポリメラーゼはRNAからRNAを合成します。
RNAウイルスの自己増殖やRNA合成の仕組みを利用すると、ワクチンもウイルスのように体内で増殖させる事が出来ます。言わば、自己増殖型mRNAワクチンとは殻の無いRNAウイルスのようなものなのです。最近の総説論文を参考にして、自己増殖型mRNAワクチンの仕組みを解説します。
自己増殖型RNAワクチン候補: mRNAワクチン開発のための代替プラットフォーム
Self-Amplifying RNA Vaccine Candidates: Alternative Platforms for mRNA Vaccine Development
Schmidt and Schnierle (2023) pathogens
https://www.mdpi.com/2076-0817/12/1/138
ヒト細胞内でRNAを「自己増殖」させるためには、1) RNAからRNAを複製する酵素 (RNA依存性RNAポリメラーゼ)、2) その酵素が認識する制御配列 (プロモーターなど) が必要となります。自己増殖型mRNAワクチンのデザインを理解するためには、その元となったウイルスのゲノム構造を知る必要があるでしょう。
多くの自己増殖型mRNAワクチンは、プラス鎖アルファウイルスであるベネズエラウマ脳炎ウイルス (VEEV)、シンドビスウイルス (SINV)、またはセンリキフォレストウイルス (SFV) 等のゲノムに基づいています。プラス鎖RNAはタンパクに直接翻訳され得るRNAです。ワクチンの自己増殖にはウイルス由来のRNAポリメラーゼが使われます。
アルファウイルスは、トガウイルス科に属する一本鎖のプラス鎖RNAウイルスです。アルファウイルスゲノムは11-12 kbです。ゲノムの5′にはメチルグアニル化キャップ、3′にはポリAテールを持ちます。二つのオープンリーディングフレーム (ORF: タンパクに翻訳され得る塩基配列) からなり、1つはRNA複製複合体 (nsp1~4)、もう1つはウイルスの外殻 (カプシドおよびエンベロープ) をコードしています。
自己増殖型mRNAワクチンのデザインの一つの例は以下の図のようになります。
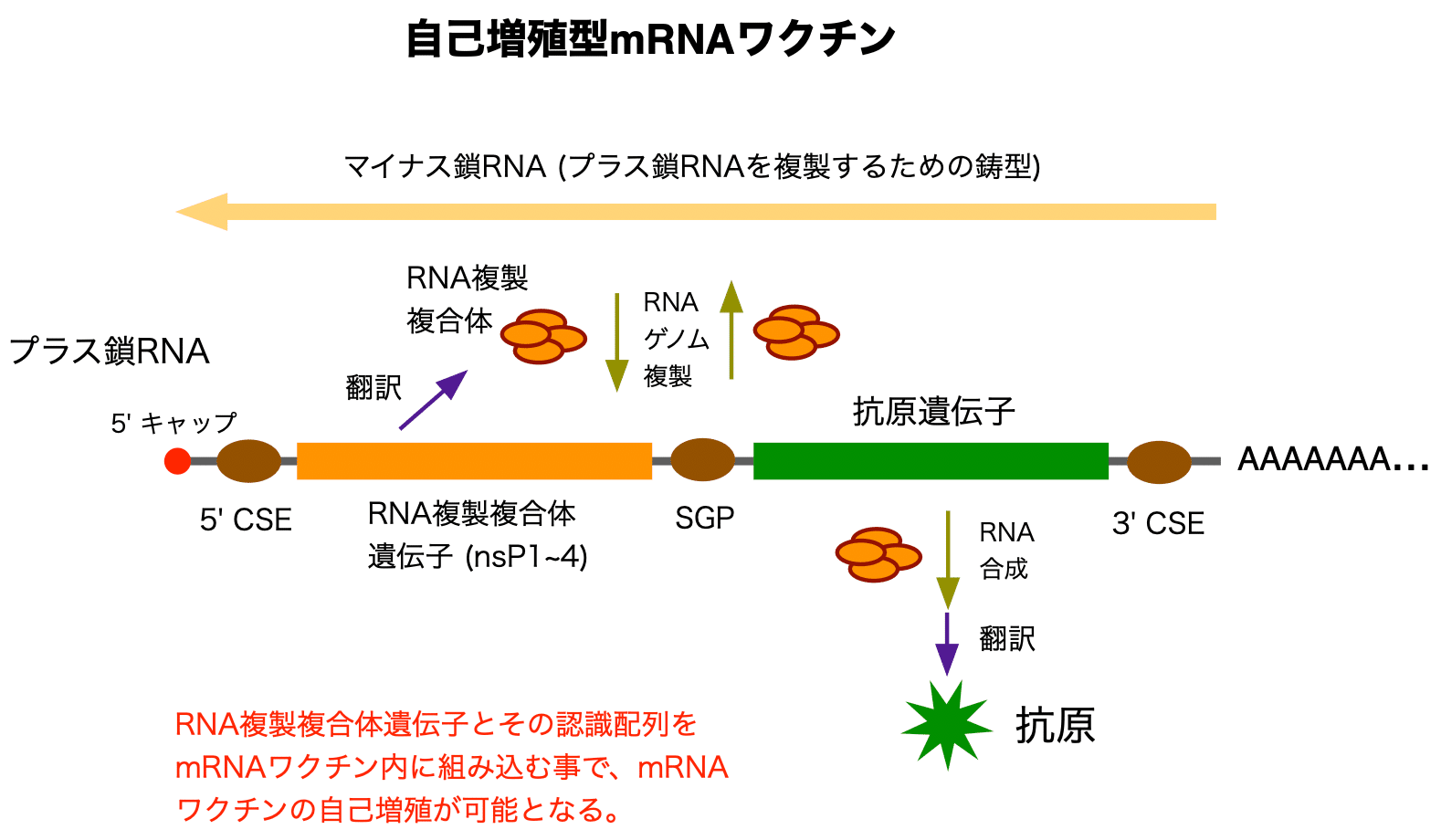
nsPはゲノムRNAの最初のORFからポリタンパク質として直接翻訳されます。4つのタンパクに分割されたnsP1~4は再び会合し、RNA複製複合体を構築します。それぞれのnsPには異なった役割があります。nsP1はキャップ酵素です。そしてnsP2はRNAヘリカーゼ兼RNAトリホスファターゼであり、また、宿主細胞のタンパク質発現の遮断を誘導します。nsP3はウイルスの複製に不可欠なウイルス-宿主、タンパク質-タンパク質相互作用を仲介します。nsP4がRNA依存RNAポリメラーゼです。
RNA複製複合体の働きは大きく分けて2つあります。1つはRNAゲノムの複製です。プラス鎖ゲノムの鋳型となる全長のマイナス鎖ゲノムを合成し、さらにそのマイナス鎖を鋳型としてプラス鎖を合成します。もう1つの働きはそれぞれの遺伝子のRNAを合成する事です。そこからRNA複製複合体や、ウイルスの外殻タンパクであるカプシドおよびエンベロープを産生します。自己増殖型mRNAワクチンのデザインは、ウイルスの外殻タンパクを抗原遺伝子で置き換えたようなものです。これはまさに「殻の無いウイルス」と呼べるでしょう。
ウイルスゲノムにはRNA合成やウイルス粒子へのパッケージングに重要な配列が含まれています。これらの要素は、シス作用型エレメント (CSE) と呼ばれます。CSEにはプロモーター機能も含まれます。二番目のORFのみのRNA合成にはサブゲノムプロモーター (SGP) が必要です。ここでは詳しく紹介しませんが、RNA複製複合体と抗原遺伝子を別々のRNAに分割する事も可能です (トランス増殖型RNAワクチン)。
mRNAワクチンの製造法には根本的な問題があり、どうしても鋳型となったDNAが混入してしまいます。そうしたDNAはゲノムにどのように作用するか分かりませんし、癌の誘発やゲノム改変に繋がるリスクがあります。そしてこれはコロナワクチンだけではなく、次世代mRNAワクチンでも同様です。自己増殖型mRNAワクチンのほぼ全体がゲノムに取り込まれ、生涯ワクチンを周囲に拡散する人が出現する可能性さえあるのです。頻度は高くないとしても、仮にそうした人が一人生まれただけでも大問題となってしまいます。
ワクチン接種者の体内でワクチンの遺伝情報が増殖するという事は、「ワクチンに感染」したとも言える状態です。私には自己増殖型mRNAワクチンの研究は「形を変えた機能獲得研究」のようにも見えます。変異速度の早いRNAウイルスの複製システムを流用している以上、自己増殖型mRNAワクチンも増殖の過程で変異します。突然変異の主な原因はゲノム複製の際のコピーミスです。
体内でRNAを複製する際にはコピーミスも起こるでしょう。そもそも全くエラーを起こさずにゲノムを複製する仕組みを作る事など出来ないのです。DNAゲノム複製の場合は相同性組換え、塩基除去修復、ヌクレオチド除去修復、ミスマッチ修復など多様なDNA修復機構がエラーを最小限に抑えますが、RNAワクチンの複製にはそうした修復機構は適用されません。突然変異による予測されない抗原の出現は自己免疫疾患を誘発する可能性があります。
進化は変異と選択によって起こるものです。進化の本質は「増えやすいものは増えやすい」と言う単純な同語反復です。自己増殖型mRNAワクチンにおいて懸念される最悪の事態の1つは、人体の中で「自己増殖型mRNAワクチンが進化」し、「増殖しやすいワクチン」や「免疫系を回避しやすいワクチン」、「感染しやすいワクチン」が派生する事です。想定される状況は、まさにワクチン接種者の「人体での人工進化による機能獲得実験」です。
これまでのmRNAワクチンはスパイクタンパク生産のブレーキのないワクチンでしたが、自己増殖するmRNAワクチンはさらに遺伝子の増殖にすらブレーキがありません。それどころかむしろ加速するアクセルが付いているようなものです。ワクチン接種者から他者にワクチンを感染させる事態が発生すると、社会の中でワクチン感染が蔓延し、ワクチン変異株が周囲に拡散されるバイオハザードのような状況すら理論的には起こり得るのです。もはやこうなるとSFの世界そのものです。しかし、これは絵空事では無いのです。現在、自己増殖型mRNAワクチンはmRNAワクチンの研究の流れの中で現実のものになっています。mRNAワクチンの研究自体が倫理的に大きな危険性を孕むものと私自身は強く感じています。